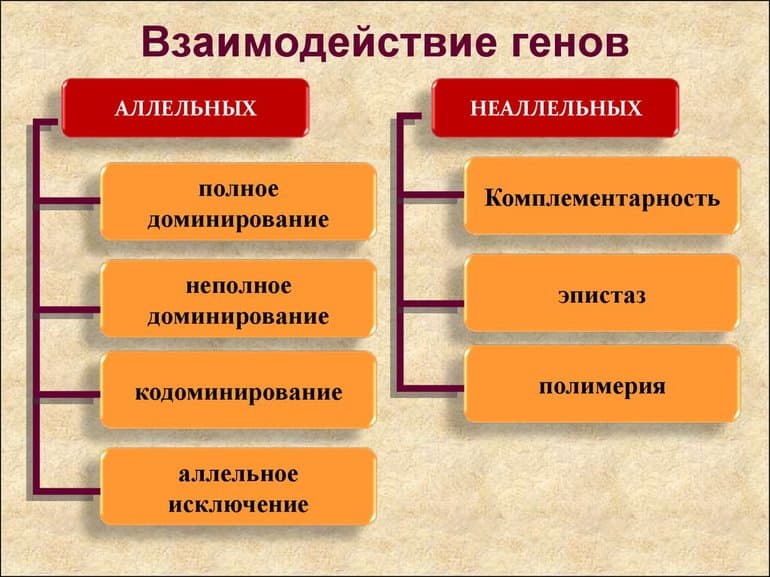
Главные закономерности наследования признаков у живых организмов были установлены Грегором Менделем. Как известно, все живые существа обладают большим количеством признаков (цвет семян и их форма у растений, тёмный или светлый окрас у животных и так далее), которые определённым образом наследуются от родительских особей. Взаимодействие аллельных генов определяет, какие признаки проявятся у потомка.
Полное доминирование
Одним из главных понятий в генетике является понятие признака — некоторой определённой особенности организма или его части, по которой особь отличается от другой. Признаком в генетическом понимании можно считать любую характеристику на уровне внешних черт, физиологических или биохимических особенностей:
- вес;
- цвет;
- рост;
- набор веществ определённого типа в органах и клетках;
- совокупность определённых белков и так далее.
Все признаки одного организма называются фенотипом. Мендель осознал, что все характеристики, которые можно использовать в генетическом анализе, должны выявляться постоянно (независимо от возраста, внешних условий и другого) и должны иметь два или более альтернативных проявлений.
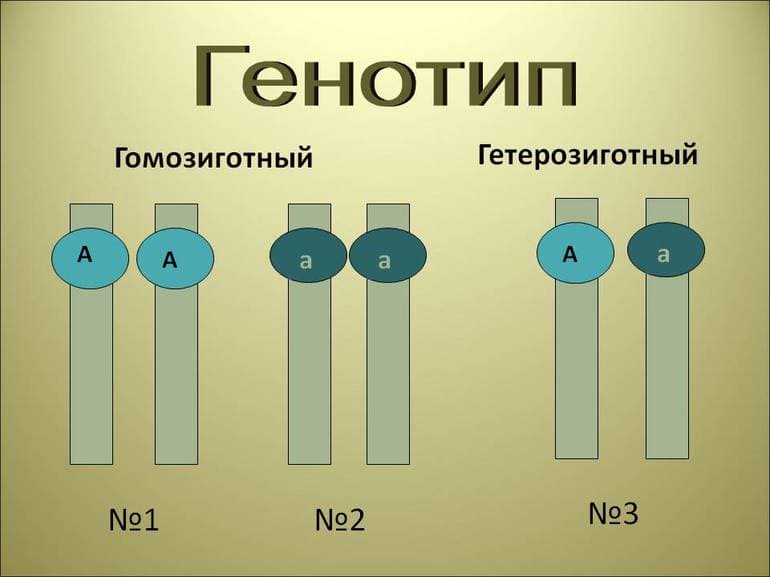
Аллельные гены (аллели) — это гены, отвечающие за один признак, но в разных его проявлениях (например, цвет семян — это один признак, который кодирует один ген, но проявляться может или жёлтый, или зелёный в зависимости от того, какой аллель является доминирующим).
Доминантные и рецессивные аллели одного гена обозначают, как правило, одинаковыми большой и маленькой буквой соответственно. Потомство от скрещивания двух особей с разными альтернативными проявлениями называют гибридным, а отдельного представителя такого потомства — гибридом.
Скрещивание, в котором проводится анализ одной пары альтернативных характеристик, называется моногибридным. Например, цвет семян гороха определяется геном sgr. Нормальный (доминантный) аллель этого гена будет обозначаться как А (он даёт функциональный продукт, который обуславливает разрушение хлорофилла и, соответственно, жёлтый цвет семян), а мутантный рецессивный аллель (ген не даёт функционального продукта, и семена остаются зелёными) как а. Именно поэтому такая мутация, когда теряется способность кодировать полноценный продукт, и обуславливает появление рецессивной формы этого гена.
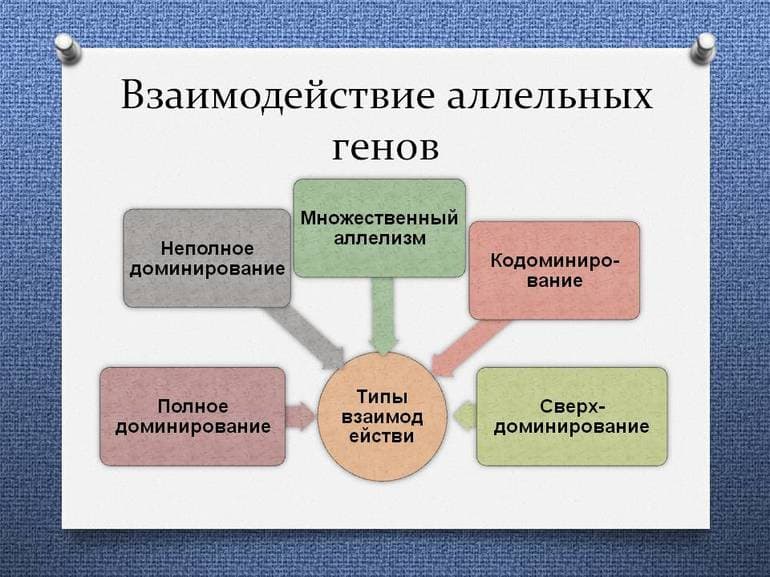
Неполное доминирование
При полном доминировании и сверхдоминировании проявление признака не зависит от количества (два или один) доминантных аллелей в генотипе. Тем не менее в природе часто встречаются ещё и другие типы взаимодействия аллельных генов. Причина может состоять в том, что рецессивный аллель не даёт функционального продукта, но характеристика зависит от количества копий доминантного аллеля в генотипе. Например, окрас цветов в красный цвет у львиного зева Antirrhinum majus зависит от количества красного пигмента, а он образуется вследствие активности фермента, который контролируется определённым геном:

- два гена АА обуславливают интенсивный красный окрас;
- гомозигота аа (отсутствие фермента и, соответственно, пигмента) характеризуется белым окрасом цветков;
- гетерозигота имеет в два раза меньшую интенсивность окраса в отличие от гомозиготы по доминантному аллелю (в два раза меньше фермента и пигмента) — цветы розового цвета.
Аналогично наследуется кучерявость перьев голубей: особь, гетерозиготная по гену кучерявости, имеет волнистые перья. Такой тип взаимодействия аллельных генов называется неполным доминированием. При неполном доминировании гетерозигота характеризуется фенотипом, отличным от обеих гомозигот — с этой поправкой первый закон Менделя остаётся без изменений. Что касается результатов скрещивания гетерозигот, то расщепление по генотипу и фенотипу будут совпадать и составлять 1:2:1.
Кодоминирование и множественный аллелизм

Аналогичные расщепления характерны и для кодоминирования с той разницей, что потомки от скрещивания гомозигот разных типов будут характеризоваться наличием фенотипических признаков обоих родителей одновременно. То есть в этом случае оба аллеля являются доминантными: образуют функциональные продукты, которые немного отличаются по некоторым характеристикам. Такая ситуация довольно часто наблюдается для ферментов, когда два аллеля обуславливают синтез двух белков — изоферментов — с практически одинаковой активностью, но при этом они отличаются молекулярной массой (один из белков имеет дополнительный структурный домен и так далее).
Эти примеры требуют важного уточнения. Поскольку в двух гомологичных хромосомах могут находиться только два разных аллеля одного гена, ещё не означает, что в группе особей ген обязательно имеет два аллеля. На самом деле, в большинстве случаев гены существуют в виде нескольких (теоретически неограниченное количество) разных аллелей. Это явление получило название множественный аллелизм.
Одним из примеров множественного аллелизма является система групп крови АВ0. Хорошо известны четыре группы крови — 0, А, В и АВ — определяются тремя аллелями одного гена: ІА, ІВ, і0. Аллели ІА и ІВ являются кодоминантными (отвечают за формирование на поверхности эритроцитов антигенов двух типов — А и В), но при этом доминируют над аллелем і0, который не продуцирует ни одного антигена.
При наличии трёх аллелей возможно шесть комбинаций между ними, которые можно получить, используя таблицу Пеннета. Но рецессивность і0 сводит количество фенотипов (групп крови) к четырём: группе 0 соответствует генотип і0і0, группе А — генотипы ІАІА и ІАі0, группе В — ІВІВ и ІВі0, группе АВ — генотип ІАІВ.
Причины отклонений от менделевских расщеплений
Менделевские расщепления в принципе могут быть реализованы только для видов живых организмов с нормальным половым процессом, диплоидностью обоих полов и при условии отсутствия нарушений в мейозе. Но и при выполнении указанных условий случаи отклонения от менделевских расщеплений достаточно многочисленны. При этом каких-либо исключений нет, а условия менделирования не нарушаются — просто на них накладываются другие эффекты.

Три таких эффекта — взаимодействие неаллельных генов (комплементация, кумулятивная и некумулятивная полимерия), сцепление групп генов в одной хромосоме и расположение гена в половых хромосомах. Среди других причин выделяются такие:

На самом деле, в биологии все эти эффекты могут быть сведены к взаимодействию генов в сложной системе функционирования генома, хотя не всегда такое взаимодействие можно легко описать: часто нужно брать во внимание слишком много отдельных элементов, которые влияют один на другого.